Steady States and Global Dynamics of Electrical Activity in the Cerebral CortexPhys Rev E Volume 58, Issue 3, pp 3557-3571, 1998Doi: 10.1103/PhysRevE.58.3557 School of Physics, University of Sydney, NSW 2006, Australia
C. J. Rennie
J. J. Wright
P. D. Bourke PACS numbers: 87.22.Jb, 87.22.As, 87.10+e
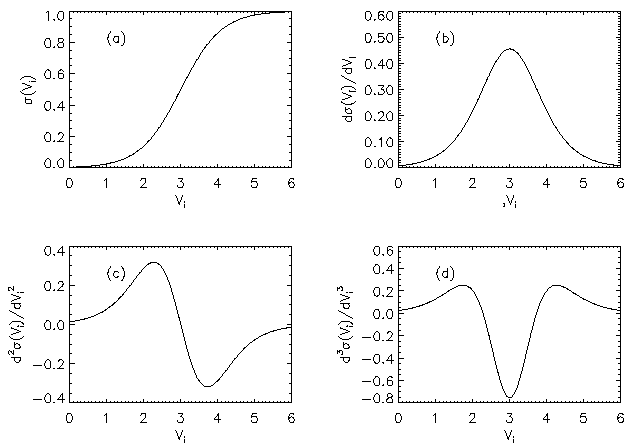
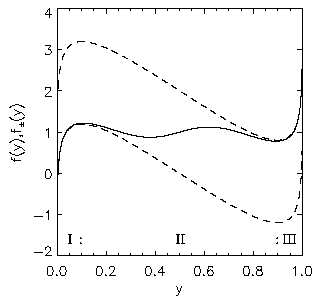
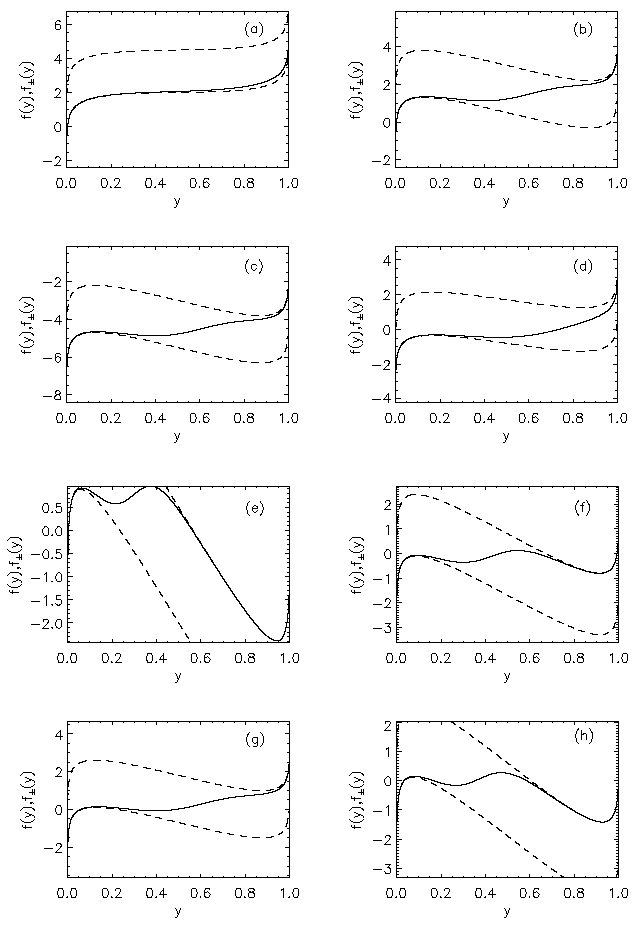
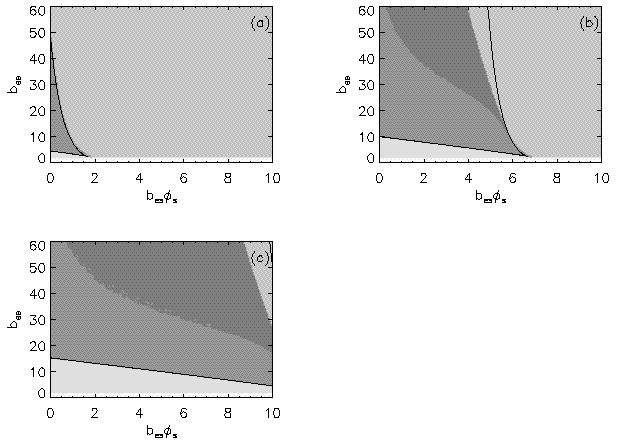
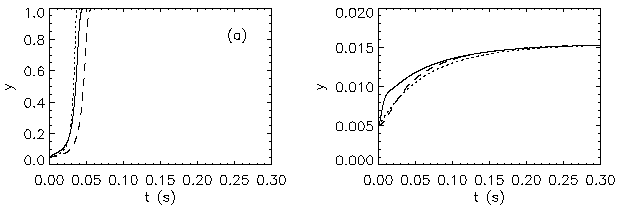
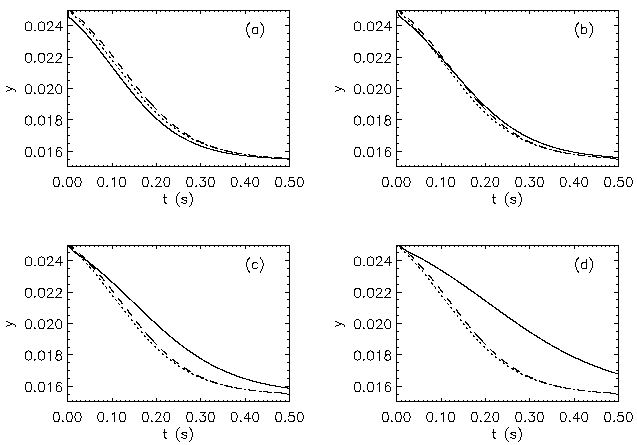
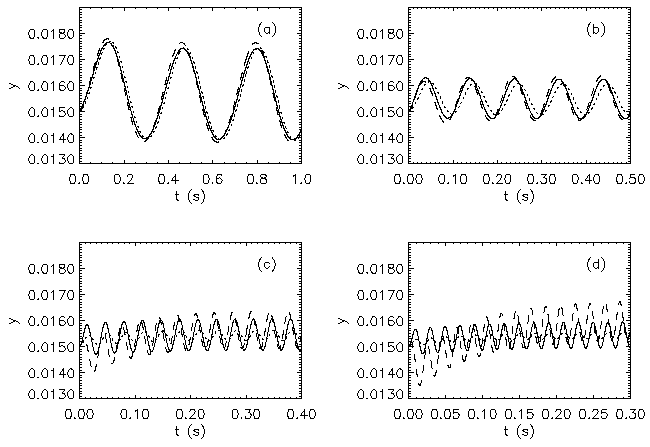
AbstractSteady states and global dynamics of electrical activity in the cerebral cortex are investigated within the framework of a recent continuum model. It is shown that, for a particular physiologically realistic class of models, at most three steady states can occur, two of which are stable. The global dynamics of spatially uniform activity states is studied and it is shown that, in a physiologically realistic class of models, the adiabatic dynamics is governed by a second-order differential equation equivalent to that for the motion of a Newtonian particle in a potential in the presence of friction. This result is used to derive a simplified dynamical equation in the friction-dominated limit. Solutions of these equations are compared with those of the full global dynamics equations and it is found that they are adequate for time-scales longer than approximately 100 ms provided dendritic integration times are less than approximately l0 ms. IntroductionRecently we developed a continuum model for the propagation of electrical activity in the cerebral cortex [1]. This model traced the evolution of quantities such as the neuronal firing rate, averaged over volumes large enough to contain many neurons, as in several previous works [2-9]. Both excitatory and inhibitory neuronal populations were included, as were the effects of nonlinear neural responses, temporal integration in the dendrites and propagation time delays in the axons. This model allowed us to write down equations for dynamics, steady state solutions, and dispersion and stability of linear waves. The above model did not include the effects of feedback on the basic parameters of the cortex, such as the threshold potential for neuronal firing, and the effective strengths of coupling between various neuronal populations. Such feedbacks are known to be of central importance in the dynamics of the actual cortex: the state of arousal (e.g., relaxed vs. alert) strongly affects the cortical response to stimuli as measured by electroencephalograms (EEGs), for example [10]. Such responses are normally termed evoked response potentials (ERPs). These responses depend not only upon fast cortical responses on time scales well below 100 ms (e.g., voltage-dependent changes in ion conductivities of neuronal membranes), but on feedbacks that evolve over longer intervals (e.g., the action of chemical neurotransmitters) [11]. Another area in which feedbacks are of importance is in the onset and termination of seizures. It is likely that the cortex operates in a state close to marginal stability [1] so as to be stable, but not so stable that all interesting behaviour is suppressed. Normal subjects are not prone to spontaneous seizures, whereas in epilepsy, for example, the cortex can undergo a transition to a seizure state in which all the neurons are firing at far above their normal rate. Such seizures are not permanent-some feedback mechanism acts to return the brain to its normal state after seconds or minutes [6,10]. The purpose of the present work is to understand better the steady states and dynamics of a generalized version of the model introduced in our previous work [1]. In Sec. II we study the possible steady-state solutions of the generalized model, deriving a steady-state equation and limits on the maximum number of steady states in certain cases. These steady states, and their basins of attraction, determine the qualitative dynamics in the absence of feedback, and provide strong constraints on the dynamics more generally for slow feedbacks. In Sec. III we study how the steady states and their interrelationships depend on the underlying parameters of the model, demonstrating that a class of models that is particularly plausible physiologically has simple properties in this respect, possessing only three steady states. Many observations of electrical activity in the cortex are made at relatively coarse spatial scales. EEGs, for example, often use scalp electrodes separated by several cm, while most short-scale features are filtered out as a result of the conductivity of the cerebrospinal fluid, skull, and scalp in any case [6]. Hence, in Sec. IV, we study the large-scale dynamics of the cortex. Furthermore, we do this in a simplified way by specializing to adiabatic dynamics on time scales much larger than the dendritic integration time, which is of order 5 - 10 ms [1,4]. We show that the spatially uniform global dynamics of the cortex can be described by an equation that is equivalent to that of a Newtonian particle moving under the influence of friction in a potential whose minimums define the stable steady states of the system. When friction dominates it is also possible to write down a simplified first-order equation for the adiabatic dynamics in the governing potential. Numerical results show that these adiabatic equations approximate the dynamics well for time scales exceeding ~100 ms. They will enable the qualitative dynamical effects of various types of feedback to be determined by looking at their effects on the potential and the resulting "forces" on the system. This ability is relevant to any analysis of feedback processes that adiabatically modify the basins of attraction of the steady-state solutions of the dynamical equations. Faster feedback mechanisms also interact with the basins of attraction, but an adiabatic approximation is not possible.
References[1] P.A. Robinson, C.J. Rennie, and J.J. Wright, Phys. Rev. E, 56, 826 (1997).[2] F.H. Lopes da Silva, A. Hoeks, A. Smits, and L.H. Zetterberg, Kybernetik, 15, 27 (1974). [3] A. van Rotterdam, F.H. Lopes da Silva, J. van den Ende, M.A. Viergever, and A. J. Hermans, Bull. Meth. Biol. 44, 283 (1982). [4] W.J. Freeman, in Induced Rhythms of the Brain, edited by E. Basar and T.A. Bullock (Birkhauser, Basel, 1991). [5] P.L. Nunez IEEE Trans. Biomed. Eng. BME-21, 473 (1974). [6] P.L. Nunez, in Neocortical Dynamics and Human EEG Rhythms, edited by P.L. Nunez (Oxford, Oxford, 1995). [7] J.J. Wright, and D.T.J. Liley, Network: Comp. Neural Syst. 5, 191 (1994). [8] J.J. Wright, and D.T.J. Liley, Biol. Cybern. 72, 347 (1995) a [9] J.J. Wright, and D.T.J. Liley, Behav. Brain Sci. 19, 285 (1996). [10] M. R. Rosenzweig, and A . L. Leiman, Physiologacal Psychology (Random House, New York, 1989). [11] 1. Segev, in Handbook of Brain Theory and Neural Networks, edited by M.A. Arbib (MIT, Cambridge, MA, 1995). [12] P.A. Robinson, J.J. Wright, and C.J. Rennie, Phys. Rev. E, in press. [13] C.J. Rennie, P.A. Robinson, and J.J. Wright, Phys. Rev. E, to be submitted. [14] D.T.J. Liley, and J.J. Wright, Network, 5, 175 (1994).
| ||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}